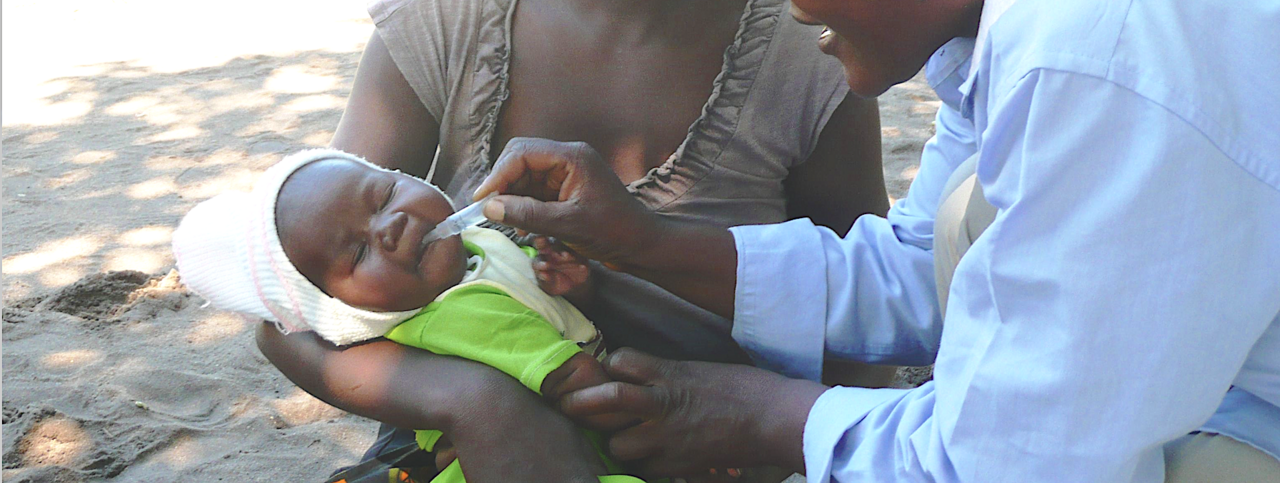

Diarrhoea and vomiting due to Rotavirus infections is a disease that will affect almost every child in the world, but mortality disproportionately affects children in low-income regions such as Africa and South East Asia. The introduction of Rotavirus vaccines in 2006 together with improved sanitation have reduced mortality from half a million to 130000 1,2. Over 120 countries have now adopted routine Rotavirus vaccination into their infant immunisation schedules. Enteric vaccine development has concentrated on live attenuated oral vaccines, consisting of live virus that no longer causes disease but is able to replicate in the gut of the host and produce a strong local immune response at the gut mucosa. The oral administration is also well suited for resource poor settings 3.
The reasons for underperformance of enteric vaccines in these settings are not well understood but are likely to be multifactorial. In the Rotavirus Vaccine Immunogenicity study (RoVI study) we set out to systematically investigate several factors that are likely to contribute to this problem. We recruited and followed up three mother-infant cohorts of participants at two resource limited locations that experience low vaccine effectiveness: Vellore in India and Blantyre in Malawi, as well as a high-income, high vaccine effectiveness control cohort in Liverpool, UK. In total we followed 664 mother-infant pairs throughout the first three months of the infants’ live, spanning the newborn immunisation schedule. All infants in the three countries received the same two dose oral rotavirus vaccine, which is licensed globally.
We first confirmed that the gap in vaccine immunogenicity existed between our study cohorts using the internationally accepted measure of seroconversion, whereby an anti-Rotavirus (anti-RV) antibody concentration is reached post-vaccination which protects the infant from serious Rotavirus (RV) disease. Our data showed that 28% of infants in the India cohort, 23% infants in Malawi cohort, and 53% infants in the UK cohort seroconverted. Another form of vaccine response that is not formally validated as a measure for protection, but which we have taken into consideration in the RoVI study, is the shedding of Rotavirus Vaccine strain (ORV) in the stool of vaccinated individuals. We used this as an indirect measure of “vaccine take”; and the differences were even more striking. In the Indian cohort, we observed a high rate of infection with a neonatal RV strain in the first week of life, this was not observed in Malawi or the UK. Our data showed that this neonatal infection is likely to play an important role in how well the Indian infants responded to vaccination.
We investigated the role that maternal antibody transfer plays in the infant vaccine response in three cohorts.
Mothers pass on antibodies through the placenta before birth and in their breastmilk to protect their child from pathogens 4. Resource poor settings tend to see higher levels of RV re-infection in adults and therefore higher antibody levels in mothers. It has been shown that RV-specific secretory Immunoglobulin A (sIgA) antibodies in breastmilk can reduce the infectivity of the vaccine strain, thereby preventing a strong immune response from the infant 4–6. However, withholding breastfeeding before a dose of oral vaccine had no significant effect on the vaccine response of infants 7. It is plausible that high levels of RV-specific antibody present in the mother and transferred through the placenta could interfere with the infants’ ability to replicate the vaccine virus and on the immune response to the vaccine 6. Disentangling the effects of maternal serum from breastmilk RV antibody on whether a child seroconverted or not is challenging.
We observed that in India and Malawi those infants whose mothers had higher levels of serum RV-antibodies had poorer immune responses to rotavirus vaccine, and shed less ORV. However, despite similar levels of maternal serum antibody to those observed in Indian mothers, vaccine shedding was not inhibited in UK infants. This highlights the potential for passively acquired antibodies to confer direct protection against viral infections (including those caused by ORV) arising at the gut mucosa. It also suggests that factors affecting the infant gut mucosa are likely to be key mediators of maternal antibody interference with ORV, rather than maternal exposure as previously hypothesized.
The second factor we investigated is whether differences in infant gut microbiota development between the cohorts plays a part in oral rotavirus vaccine immunogenicity and take.
The human gut microbiota plays an important role in intestinal and immunological development and maturation 8,9. It is therefore reasonable to expect that it will also influence immune responses to oral vaccines. Enteric live-attenuated vaccines need to replicate in the intestinal tract, in direct interaction with the intestinal flora and in an environment that is influenced and shaped by it; hence the microbiota is also likely to directly and/or indirectly impact on efficient vaccine strain replication, necessary to elicit a protective local immune response at the gut mucosa.
We observed considerable geographical differences in how the gut microbiota developed as early as the first week of the infants’ life. Malawian infants showed the greatest diversity in their microbiota, although this discrepancy waned a little with increasing age, the samples geographic origin could be predicted by its composition at all timepoints, demonstrating significant and persistent geographical/population specific microbiota development patterns. In both India and Malawi, higher microbiota diversity was associated with poorer immune response to ORV. We did not observe relationships between microbiota diversity and response to ORV among infants in the UK and interestingly none of the three cohorts showed an association between shedding and microbiota diversity. We did not identify any associations between specific genera of bacteria and seroconversion, antibody levels or shedding of the vaccine strain in the RoVI study. However, the demonstration of a consistent microbiota signature of impaired oral RV response in two independent settings suggests that high early-life microbial exposure may contribute to impaired ORV efficacy. This raises the possibility that impaired oral vaccine response might be viewed as a counterpart to hyperresponsive immune states associated with low early-life microbiota diversity, such as atopic disease.
A particular issue in resource poor settings, and the third factor that we investigated, is environmental enteric disease (EED).
EED is a subclinical disorder that occurs among inhabitants of environments with poor sanitation and hygiene. Chronic exposure to faecal pathogens is thought to cause intestinal inflammation and structural changes in the small bowel. EED is marked by increased intestinal permeability and malabsorption and has been cited as a possible cause for enteric vaccine failure 10,11. The gold standard for diagnosing this condition would be via a colonoscopy and biopsy, but for obvious reasons this cannot be carried out routinely on healthy infants. We therefore rely on measuring biomarkers for gut and systemic inflammation and gut permeability. Despite significant geographic differences observed between the cohorts with the highest levels of these markers observed in Malawi and the lowest in the UK, none of them were associated with seroconversion, RV-IgA levels or ORV shedding in any of the cohorts. This may suggest that these markers alone may not correlate or describe with sufficient sensitivity and accuracy the complex changes associated with EDD.
Conclusions from the RoVI study
For a specific factor to be classified as contributing towards enteric vaccine failure we would expect to have observed it in multiple geographic settings that show poor vaccine response, this has not been the case in this study.
Instead, we highlight the complexity of assigning RV-antibody levels alone as a measure of protection and advocate for the use of other measures such as vaccine virus shedding as an additional measure of response to better capture the disparate oral vaccine response in different populations.
We did not observe any broad associations between enteric vaccine response and specific bacteria across our cohorts. Our findings are also contrary to established descriptions of what constitutes a ‘healthy’ microbiota. Low microbiota diversity is often used as a marker of ‘dysbiosis’, but this is largely informed by studies in high-income settings, and often in adults. Our observation of enriched microbiota diversity in vaccine non-responders suggests a more complex picture, whereby the signatures of healthy microbiota vary not only by geographic setting and age, but by the health outcome under consideration.
Our study also showed that asymptomatic neonatal exposure to the highly prevalent rotavirus "neonatal strain" in Indian infants correlated significantly with increased ORV immunogenicity. This finding suggests vaccination in the neonatal period, could significantly improve ORV effectiveness in low-income countries. These data advocate for the need to investigate neonatal immunization, whether with a neonatal strain or a vaccine attenuated strain, as a pragmatic approach to achieve greater impact through modification of current rotavirus vaccination programs.
References
- Troeger, C. et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea Among Children Younger Than 5 Years. JAMA Pediatr. 172, 958–965 (2018).
- Tate, J. E. et al. 2008 estimate of worldwide rotavirus-associated mortality in children younger than 5 years before the introduction of universal rotavirus vaccination programmes: a systematic review and meta-analysis. Lancet Infect. Dis. 12, 136–141 (2012).
- Kosek, M. et al. Fecal Markers of Intestinal Inflammation and Permeability Associated with the Subsequent Acquisition of Linear Growth Deficits in Infants. Am. J. Trop. Med. Hyg. 88, 390–396 (2013).
- Chan, J. et al. Maternal antibodies to rotavirus: could they interfere with live rotavirus vaccines in developing countries? Vaccine 29, 1242–1247 (2011).
- Patel, M., Steele, A. D. & Parashar, U. D. Influence of oral polio vaccines on performance of the monovalent and pentavalent rotavirus vaccines. Vaccine 30 Suppl 1, A30-35 (2012).
- Moon, S.-S. et al. Inhibitory effect of breast milk on infectivity of live oral rotavirus vaccines. Pediatr. Infect. Dis. J. 29, 919–923 (2010).
- Rongsen-Chandola, T. et al. Effect of withholding breastfeeding on the immune response to a live oral rotavirus vaccine in North Indian infants. Vaccine 32 Suppl 1, A134-139 (2014).
- Nicholson, J. K. et al. Host-gut microbiota metabolic interactions. Science 336, 1262–1267 (2012).
- Hooper, L. V., Littman, D. R. & Macpherson, A. J. Interactions between the microbiota and the immune system. Science 336, 1268–1273 (2012).
- Korpe, P. S. & Petri, W. A. Environmental enteropathy: critical implications of a poorly understood condition. Trends Mol. Med. 18, 328–336 (2012).
- Church, J. A. et al. Exploring the relationship between environmental enteric dysfunction and oral vaccine responses. Future Microbiol. 13, 1055–1070 (2018).




Please sign in or register for FREE
If you are a registered user on Research Communities by Springer Nature, please sign in